دایناسورها در میان خزندگاندو تبار از تبارهای خزندگان که در پایان دوران پالئوزوئیک تکامل یافته بودند، آرکوسورومورفها۱۴ و لپیدوسورومورفها۱۵ بودند. انشعاب این دو گروه از نیای مشترکشان در اواخر پالئوزوئیک رخ داد. مهمترین تفاوتهای این دو گروه در شیوه قرارگرفتن پاها بود. پاهای لپیدوسورومورفها مشابه وضعیتی که در بسیاری از مهرهداران خشکیزی دوران پالئوزوئیک دیده میشد، به سمت دو طرف بدن خارج میشد، اما پاهای آرکوسورومورفها با وضعیتی تازه در زیر بدن و عمود بر سطح زمین قرار میگرفت. هرکدام از این دو گروه شامل زیرگروههای مختلفی است که بهتدریج طی دوران مزوزوئیک (۲٥۲ تا ٦٦ میلیون سال پیش) تکامل یافتند. مارها، مارمولکها و احتمالاً لاکپشتها چندتا از تبارهایی هستند که در دل تبار لپیدوسورومورفها تکامل پیدا کردند. تمساحها و تروسورها (خزندگان پروازگر) و دایناسورها هم چند تا از تبارهایی هستند که در دل تبار آرکوسورومورفها تکامل یافتند. خزندگان دریازی هم (که به غلط دایناسور دریازی نامیده میشوند) احتمالاً از خویشاوندان دور لاکپشتها و جزء تبار لپیدوسورومورفها بودهاند. همه این گروهها در دوران مزوزوئیک تنوع گستردهای داشتند؛ مثلاً، خویشاوندان تمساحها در اوایل دوران مزوزوئیک تنوع ریختی زیادی داشتند و شامل انواع مختلفی از خزندگان کوچک و بزرگ خشکیزی دونده، نیمهآبزی یا کاملا دریازی میشدند. حتی گروههایی از تمساحها گیاهخوار بودند. برخی از آنها دندانهای خود را از دست داده، مثل لاکپشتها و پرندگان منقار داشتند و حتی برخی از این تمساحهای گیاهخوار بیدندان، روی دوپای عقب خود راه میرفتند و دوپا شده بودند که نباید این موجودات را با دایناسورها اشتباه گرفت. دایناسورها هم مثل تمساحها آرکوسورومورف هستند، اما تباری دیگر را تشکیل میدهند.
مقدمهتوصیف آرکئوپتریکس۱ در سال ۱۸۶۱ چنان تأثیرگذار بود که داروین در ویرایش چهارم کتاب «خاستگاه گونهها» از آن نام برد و به ویژگیهای مهمی اشاره کرد که آرکئوپتریکس را به «حلقه گمشده» میان خزندگان و پرندگان تبدیل میکنند: «...این مرغ غریب با دُمی دراز که در هر بند آن جفتی پر روئیده و بالهایی آراسته به دو ناخن آزاد [از هم] در الواح آهکی سولنهافن پیدا شده است. قویاً هیچ کشف متأخر دیگری چنین به قوت نشان نمیدهد که پیش از این، چه اندازه اندک درباره باشندگان سابق زمین میدانستیم» (داروین، خاستگاه گونهها، ۱۸۶۶، ص .۳۶۷).
اکنون مدتها از آن زمان میگذرد. اطلاعات بسیار زیادی درباره باشندگان سابق زمین، به خصوص نیاکان پرندگان، به دست آمده است، اما هنوز هم کل این اطلاعات چیزی نیست جز نقاطی که باید به هم متصل شوند. گرچه این نقاط ماهیتی حقیقی و عینی دارند، خطوطی که میان نقاط یافتشده رسم خواهیم کرد، فرضیات و نظریاتی در علم تجربی هستند که باید با نگاهی ابطالگرا و سنجشگرانه مورد قضاوت قرار گیرند؛ مثلاً، فرضیاتی برای پاسخ به این پرسشها که نخستینبار پرواز چگونه تکامل یافت؟ پرها برای پرواز تکامل یافتند یا پرواز معلول تکامل پرها بود؟ نخستین پرندگان از زمین به هوا میپریدند یا از شاخههای درختان؟ شاید هرگز نتوانیم قطعاً این فرضیات و نظریات را قبول یا رد کنیم، اما با کشف سنگوارههای بیشتر و پیوستهتر شدن نقاط شناختهشده، به حقیقت اندکی نزدیکتر میشویم.
کشف رابطه پرندگان و دایناسورهاداروین در ویرایشهای نهایی کتابش، به نظریات دوستش، تامس هاکسلی۲ ، درباره آرکئوپتریکس نیز اشاره میکند. هاکسلی در خلال سالهای ۱۸٦۸ تا ۱۸۷۰ در مقالاتی۳ به شباهتهای آرکئوپتریکس (و پرندگان امروزی) با دایناسورها اشاره کرد و پرندگان امروزی را نسل بازمانده از تکامل دایناسورها دانست. تا اوایل سده بیستم نیز دیرینهشناسان دیگر مانند هاکسلی فکر میکردند۴ و البته ما هم امروزه همانند هاکسلی فکر میکنیم. از میان ۳٥ صفت مشترکی که هاکسلی میان پرندگان و دایناسورها نشان داد، هفده مورد هنوز هم درست و دقیق شمرده میشوند. با وجود این، پس از مرگ هاکسلی، فرضیه او به فراموشی سپرده شد.۵ در همان سال ۱۸٥۹ که کتاب خاستگاه گونهها برای نخستینبار منتشر شد، کودکی در دانمارک متولد شد که در میانسالی فرضیه هاکسلی را زیر سوال برد. گرهارد هیلمن۶، پزشک، تصویرگر و دیرینهشناس دانمارکی، در سال ۱۹۱٦ کتابی به زبان دانمارکی نوشت به نام «خاستگاه پرندگان»۷ که ده سال بعد به انگلیسی ترجمه شد. او در این کتاب به روشنی نشان داد که پرندگان بیش از هر گروه دیگر به دایناسورها شباهت دارند، اما رابطه میان دایناسورها و پرندگان را رد کرد؛ به این علت که عقیده داشت دایناسورها فاقد استخوان ترقوه۸ هستند؛ درحالیکه پرندگان ترقوه دارند. استخوانهای ترقوه پرندگان از دو سو به هم متصل شده و استخوانی ۷ مانند تشکیل دادهاند (موسوم به furcula یا چنگالک که در فارسی به غلط «جناغ» نامیده میشود). هیلمن با دست گذاشتن بر همین موضوع فرضیه هاکسلی را رد کرد و سررشته تکامل پرندگان برای هفتاد سال از دست دیرینهشناسان گم شد.
هیلمن اشتباه میکرد. در سال ۱۹۲٤ سنگواره دایناسوری در مغولستان کشف و توصیف شد که درست شبیه پرندگان، استخوان ترقوهای ۷ مانند داشت.۹ در سال ۱۹۳٦ سنگواره دایناسور دیگری در آمریکای شمالی کشف شد که باز هم استخوان ترقوه آشکاری داشت. برای دیرینهشناسان امروز، نمونههای متعدد دیگری از دایناسورهایی با استخوان ترقوه، هر تردیدی را درباره خویشاوندی پرندگان و دایناسورها از میان برده است.۱۰ اما تا سال ۱۹۸٦ که ژاک گوتیه۱۱ برای نخستینبار با تحلیلهای فیلوژنتیک نشان داد آرکئوپتریکس و پرندگان از تبار دایناسورها هستند، فرضیه هاکسلی همچنان در محاق بود.۱۲ پس از انتشار تحلیلهای فیلوژنتیک گوتیه، صدها پژوهش مشابه با روشهای مختلف به تحلیل فیلوژنتیک دایناسورها پرداخته و همگی نشان دادهاند که حق با گوتیه (و هاکسلی) بوده است و پرندگان دایناسور هستند.۱۳
 شکل ۱: نخستین سنگواره آرکئوپتریکس مشهور به نمونه موزه لندن (سمت راست) کهفاقد قسمت اعظم بقایای سر و گردن است
شکل ۱: نخستین سنگواره آرکئوپتریکس مشهور به نمونه موزه لندن (سمت راست) کهفاقد قسمت اعظم بقایای سر و گردن است
و مشهورترین نمونه آرکئوپتریکس موسومبه نمونه موزه برلین (سمت چپ) که تصویر کاملی از سر و دندانهای ریز در آندیده میشود.
دایناسورها چه جانورانی بودند؟برخی فکر میکنند هر خزنده غولپیکر، یا هر خزنده ماقبل تاریخ یا حتی همه موجودات ماقبل تاریخ دایناسور هستند. هیچ کدام از اینها درست نیست. خزندگان تا پایان دوران پالئوزوئیک (۲٥۲ میلیون سال پیش) به گروههای متعددی منشعب شده بودند که امروز آنها را از سنگوارههایشان میشناسیم و ردهبندی میکنیم. آن زمان هنوز هیچ کدام از گروههای مشهور خزندگان، مثل مارمولکها و مارها و لاکپشتها و تمساحها وجود نداشتند، اما تبارهایی تکامل یافته بودند که بعدها منتهی به تکامل گروههای متعارف امروزی خزندگان شدند.
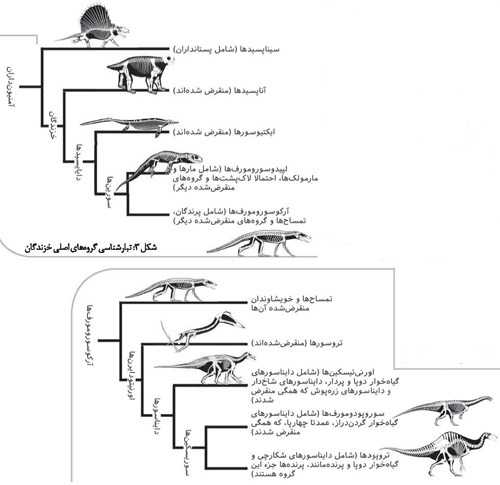 شکل ۲: استخوان ترقوه یا چنگالک در آرکئوپتریکس (بالا) در مقایسه با باتیرانوسورِس (پایین سمت راست) و ولاسیراپتور (پایین سمت چپ).
شکل ۲: استخوان ترقوه یا چنگالک در آرکئوپتریکس (بالا) در مقایسه با باتیرانوسورِس (پایین سمت راست) و ولاسیراپتور (پایین سمت چپ).
این استخوان ۷مانند که با رنگ روشن مشخص شده، میان دو نیمه کمربند شانهای قرار گرفته،یعنی در سمت پیشین استخوان جناغ قرار دارد و از هر طرف به استخوان غرابی وکتف میرسد.
دایناسورها در میان خزندگاندو تبار از تبارهای خزندگان که در پایان دوران پالئوزوئیک تکامل یافته بودند، آرکوسورومورفها۱۴ و لپیدوسورومورفها۱۵ بودند. انشعاب این دو گروه از نیای مشترکشان در اواخر پالئوزوئیک رخ داد. مهمترین تفاوتهای این دو گروه در شیوه قرارگرفتن پاها بود. پاهای لپیدوسورومورفها مشابه وضعیتی که در بسیاری از مهرهداران خشکیزی دوران پالئوزوئیک دیده میشد، به سمت دو طرف بدن خارج میشد، اما پاهای آرکوسورومورفها با وضعیتی تازه در زیر بدن و عمود بر سطح زمین قرار میگرفت. هرکدام از این دو گروه شامل زیرگروههای مختلفی است که بهتدریج طی دوران مزوزوئیک (۲٥۲ تا ٦٦ میلیون سال پیش) تکامل یافتند. مارها، مارمولکها و احتمالاً لاکپشتها چندتا از تبارهایی هستند که در دل تبار لپیدوسورومورفها تکامل پیدا کردند. تمساحها و تروسورها (خزندگان پروازگر) و دایناسورها هم چند تا از تبارهایی هستند که در دل تبار آرکوسورومورفها تکامل یافتند. خزندگان دریازی هم (که به غلط دایناسور دریازی نامیده میشوند) احتمالاً از خویشاوندان دور لاکپشتها و جزء تبار لپیدوسورومورفها بودهاند. همه این گروهها در دوران مزوزوئیک تنوع گستردهای داشتند؛ مثلاً، خویشاوندان تمساحها در اوایل دوران مزوزوئیک تنوع ریختی زیادی داشتند و شامل انواع مختلفی از خزندگان کوچک و بزرگ خشکیزی دونده، نیمهآبزی یا کاملا دریازی میشدند. حتی گروههایی از تمساحها گیاهخوار بودند. برخی از آنها دندانهای خود را از دست داده، مثل لاکپشتها و پرندگان منقار داشتند و حتی برخی از این تمساحهای گیاهخوار بیدندان، روی دوپای عقب خود راه میرفتند و دوپا شده بودند که نباید این موجودات را با دایناسورها اشتباه گرفت. دایناسورها هم مثل تمساحها آرکوسورومورف هستند، اما تباری دیگر را تشکیل میدهند.
خزندگان دوپااجداد دایناسورها آرکوسورهایی دوپا بودند، اما کالبدشناسی پای آنها تفاوتهای زیادی با پای تمساحها (حتی با تمساحهای دوپای گیاهخوار) داشت. گرچه هر دو گروه تمساحها و دایناسورها در اوایل دوران مزوزوئیک (۲۲۰ میلیون سال پیش) تکامل یافتند و هر دو گروه شامل انواعی دونده و دوپا میشد، اما تمساحهای دوپا تا حدود ۲۰۰ میلیون سال پیش دوام آوردند و سرانجام در رقابت بر سر شیوه زندگی از دایناسورهای دوپا شکست خوردند. از حدود ۲۰۰ میلیون سال پیش گروههایی از دایناسورها هم رفتهرفته به دلیل بزرگشدن جثه دوباره به حالت چهارپا درآمدند.
 شکل٥. آرکئوپتریکس شاید تنها به این دلیل مهمتر از سایر دایناسورهای پردارتلقی میشود که ۱٦۰ سال پیش کشف شد؛ یعنی خیلی زودتر از باقی خویشاوندانش وبرای پرهایی که در سنگوارهاش آشکارا دیده میشوند، طی همه این سالهاعنوان «نخستین پرنده» را داشت،
شکل٥. آرکئوپتریکس شاید تنها به این دلیل مهمتر از سایر دایناسورهای پردارتلقی میشود که ۱٦۰ سال پیش کشف شد؛ یعنی خیلی زودتر از باقی خویشاوندانش وبرای پرهایی که در سنگوارهاش آشکارا دیده میشوند، طی همه این سالهاعنوان «نخستین پرنده» را داشت،
اما بررسیهای اخیر نشان میدهند که اگرآرکئوپتریکس پرنده است (که هست) دایناسورهای پردار دیگر مثل ولاسیراپتورنیز باید پرنده شمرده شوند. از سوی دیگر، توانایی پرواز میلیونها سال بعداز آرکئوپتریکس تکامل یافته است و به این ترتیب، نخستین پرندگان، پروازگرانحقیقی نبودند.
خزندگان خونگرمتروسورها (خزندگان پروازگر) هم گروه دیگری از آرکوسورومورفها بودند که خویشاوندی نزدیکتری با دایناسورها داشتند. تروسورها بالهایی پوستی داشتند، شبیه بال خفاش، با این تفاوت که در خفاش پرده پوستی بال میان چهار انگشت کشیده شده، اما در تروسورها فقط یکی از انگشتان دست درازشده و حامل بال پوستی بود. دایناسورها و تروسورها نزدیکترین خویشاوندان یکدیگر بودند، اما تروسورها را نباید با دایناسورها اشتباه کرد. گردنهای کشیده، پوششی از پرهای کرکمانند و کیسههای هوایی متصل به شش (هم برای کاهش جرم حجمی بدن، هم برای بهبود کارایی تنفسی و هم برای خنککردن بدن هنگام فعالیت) ویژگی مشترک دایناسورها و تروسورها بود که در تمساحها و لپیدوسورومورفها پیدا نمیشد. وجود پرهای کرکمانند در نیای مشترک تروسورها و دایناسورها نشان میدهد هر دو گروه خونگرم بودهاند. خود دایناسورها شامل سه زیرگروه بزرگ میشوند: سوروپودمورفها که دایناسورهایی گیاهخوار با گردنهای دراز بودند. سوروپودومورفهای اولیه دوپا بودند، اما ۲۰۰ میلیون سال پیش با بزرگشدن و سنگینشدن بدن آنها، گروهی از آنها حرکت روی چهارپا را انتخاب کردند. سوروپودومورفهای چهارپا طی چند میلیون سال به بزرگترین جانورانی تبدیل شدند که بر زمین قدم برداشتند. حدس میزنیم سوروپودومورفهای کوچکجثه اولیه هم پر داشتهاند، اما با بزرگشدن جثههایشان، پرهایشان را از دست دادهاند. تروپودها عمدتاً دایناسورهای شکارچی هستند. همه تروپودها دوپا باقی ماندند، اما همه آنها شکارچی باقی نماندند. پرهای تروپودها تکامل و تنوع زیادی پیدا کرد و گروههایی از تروپودها صاحب پرهایی تزئینی و بزرگ روی دمها و دستهای خود شدند. اورنیتیسکینها گروه دیگری از دایناسورها بودند که مثل سوروپودومورفها گیاهخوار بودند و مثل تروپودها دوپا؛ نیز نمونههای پردار زیادی از آنها در سنگوارهها پیدا شده است. گروههایی از آنها هم با بزرگشدن جثه به چهارپایی رو آوردند. دایناسورهای زرهپوش، شاخدار و منقاراردکیها گروههایی از اورنیتیسکینها بزرگجثه هستند.
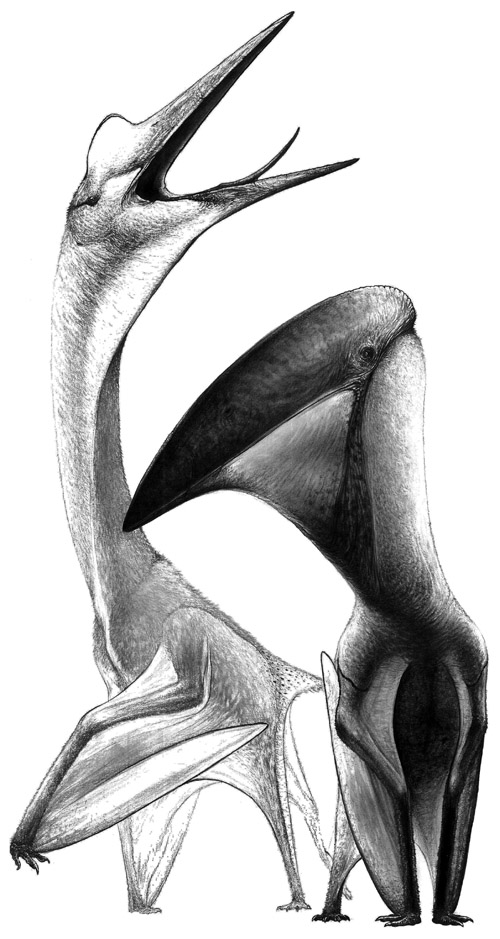 شکل ٦: نمونههایی از تروسورها که دایناسور نبودند، اما نزدیکترینخویشاوندان دایناسورها به شمار میروند. هر دو نمونه از خانواده اژدرخیدها(Azhdarchidae) و از تروسورهای کرتاسه بالایی اند که جثههایی بزرگ (عرضبال ۱۰ متر و حتی بیشتر)، دم کوتاه و منقارهایی بدون دندان، داشتند. نمونهگردنکوتاهتر Hatzegopteryx (هاتزگوپتریکس) نام دارد که در کرتاسهبالایی، در جزایری زندگی میکرد که امروزه اروپای مرکزی را میسازند و ازبچه دایناسورها تغذیه میکرد.
شکل ٦: نمونههایی از تروسورها که دایناسور نبودند، اما نزدیکترینخویشاوندان دایناسورها به شمار میروند. هر دو نمونه از خانواده اژدرخیدها(Azhdarchidae) و از تروسورهای کرتاسه بالایی اند که جثههایی بزرگ (عرضبال ۱۰ متر و حتی بیشتر)، دم کوتاه و منقارهایی بدون دندان، داشتند. نمونهگردنکوتاهتر Hatzegopteryx (هاتزگوپتریکس) نام دارد که در کرتاسهبالایی، در جزایری زندگی میکرد که امروزه اروپای مرکزی را میسازند و ازبچه دایناسورها تغذیه میکرد.
نمونه گردندراز، Quetzalcoatlus (کتزالکواتلس) نام دارد که در کرتاسه بالایی در آمریکای شمالی، لاشهخواریمیکرد. قامت کتزالکواتلس به قد زرافه بود. بدن تروسورها پوشیده از پرهایمومانند بــود (تصویرسازی امیرحسین بهجتی)
خزندگان پردار امروزیپرندگان امروزی تنها گروه دایناسورها هستند که تا امروز زنده ماندهاند. دیواره کامل قلب میان دو بطن قلب (که مانع مخلوطشدن خون تیره و روشن میشود)، سنگدان، پوسته تخم آهکی، ساخت لانه برای تخمگذاری، مراقبت از بچهها، قرارگرفتن پاها زیر بدن، وجود دندههای گردنی و بسیاری صفات کالبدشناختی دیگر، ویژگیهایی مشترک میان همه آرکوسورومورفها هستند؛ پرندگان و تمساحها (و تا جایی که از سنگوارهها میدانیم، تروسورها و دایناسورهای منقرضشده نیز) واجد این صفتها هستند. راهرفتن روی دو پا، وجود پر و شاهپر، سوراخ بودن مفصل لگن، تعدد مهرههای لگن و بسیاری از صفات کالبدشناختی دیگر نیز نشان میدهند که پرندگان جزء تروپودها هستند. داستان تکامل پرواز در پرندگان، در حقیقت داستان تکامل تروپودهاست.
نخستین پروازهاحدود ٤۰۰ میلیون سال پیش، گروهی از بندپایان ششپا که ما معمولاً آنها را حشرات مینامیم، به ترتیبی که هنوز درباره آن اطمینان نداریم (شاید از چینهای پوستی جانبی بدن یا شاید از توسعه آبششهای نیایی متصل به پاها) بال درآوردند و شروع به پرواز کردند. حشرات نخستین جانورانی بودند که توانستند با کمک اندامهایی که هوا را به هم میزدند، از زمین جدا شوند و شروع به پرواز کنند. ۲۲۸ میلیون سال پیش گروهی از خزندگان که پیشتر قلبی کاملاً چهارحفرهای، بدنی پوشیده از پرهای کرکمانند و سوختوسازی احتمالاً خونگرم داشتند، با بالهایی پوستی که میان انگشت شماره ٤ دست و پهلوها و پاها کشیده شده بود، شروع به پرواز کردند. اینها تروسورها بودند؛ نخستین مهرهداران پروازگر و تنها گروه پروازگری که ٦٦ میلیون سال پیش، نسلش کاملاً منقرض شد. حدود ۱٤٥ میلیون سال پیش، گروه دیگری از خزندگان پردار به شیوهای متفاوت با تروسورها شروع به پرواز کردند. آنها مجهز به نوع خاصی از پرهای مستحکم و بزرگ بودند و با کمک همین پرها شروع به پریدن میان شاخههای درختان کردند. اینها دایناسورها بودند و نسل گروهی از دایناسورهای پردار پروازگر تا امروز باقی مانده است. سرانجام حدود ٥۳ میلیون سال پیش، گروهی از پستانداران صاحب بالهایی پوستی شدند و شروع کردند به پرواز. این پستانداران پروازگر خفاشها هستند و تا امروز هنوز منقرض نشدهاند. درباره اینکه نخستین حشرات، نخستین تروسورها و نخستین خفاشها چطور پرواز را شروع کردند، تقریباً هیچچیز نمیدانیم. هنوز نمونهای از سنگواره اجداد این گروهها کشف نشده که بالهایی نصفه داشته باشند. قدیمیترین نمونههای کشفشده هرکدام از این گروهها هم بالهایی کامل و مناسب برای پرواز داشتهاند. اما خوشبختانه از مراحل اولیه تکامل پرواز در پرندگان، شواهد خوبی به جا مانده است.
دایناسورهای بالدارشکی نیست که پر پیش از پرواز تکامل یافته است. تا مدتها قدیمیترین موجود پرداری که میشناختیم آرکئوپتریکس بود که شاهپرهایی کامل دارد. اما طی بیست ـ سی سال اخیر، نمونههای جدیدی از سنگوارههای دایناسورهای پردار کشف شدند که به روشنی مراحل تکامل پر را نشان میدهند. دایناسورهایی که تنها پوششی از پوشپر دارند؛ دایناسورهایی که تنها پوششی از کرکپر دارند؛ دایناسورهایی که ترکیبی از پر و فلس در کنار هم روی پوست خود دارند؛ دایناسورهایی که پرهایی با شکلهای متفاوت نسبت به پرهای پرندگان امروزی دارند و سرانجام دایناسورهایی که پرهایی بزرگ اما تیغمانند دارند. علاوه بر اینها از دیرباز میدانستیم که بدن تروسورها، یعنی همان خزندگانِ پروازگرِ بالپوستی که نزدیکترین خویشاوندان دایناسورها به شمار میروند، از پوششی مومانند پوشیده شده بود. مدتها بر سر اینکه شاید این موها نیز نوعی پر ابتدایی باشند، بحث وجود داشت تا سرانجام سال گذشته سنگواره تروسوری پیدا شد که موهای روی سطح پوستش، به وضوح الگویی منشعب داشتند. این یافته کافی بود تا وجود پر را به نیای مشترک دایناسورها و تروسورها نیز تعمیم بدهیم. طبق آنچه در سنگوارهها مشهود است، نخستین پرها که در دایناسورها و تروسورها و نیای مشترک آنها وجود داشته، چیزی بوده است شبیه کرکهای زردرنگ جوجه پرندگان امروزی. اما سؤال بزرگی همچنان باقی میماند: چرا برخی دایناسورها صاحب شاهپر شدند؟
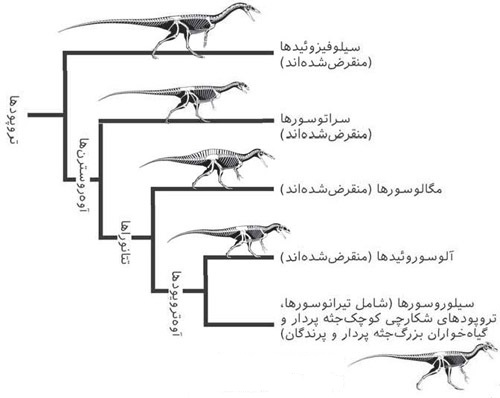 شکل ۷. تبارشناسی تروپودها و جایگاه پرندگان در میان آنها.
شکل ۷. تبارشناسی تروپودها و جایگاه پرندگان در میان آنها.
ماجرای تاژک باکتریمخالفان نظریه تکامل برای رد کردن این نظریه دست به دامن تکامل ساختارهایی از جمله تاژک باکتریها میشوند. تاژک باکتری از چندین نوع پروتئین مختلف ساخته شده که تنها در کنار همدیگر میتوانند تاژک را به پیشران یاختهای تبدیل کنند. غیبت هرکدام از این پروتئینها کافی است تا تاژک کار نکند. مخالفان نظریه تکامل میگویند که احتمال پیدایش بختانه هرکدام از این پروتئینها بسیار اندک است؛ چه رسد به اینکه حدود ۱۰ تا۲۰ پروتئین مختلف همزمان جهش یابند و طوری کنار هم قرار گیرند که بتوانند به تاژک تبدیل شوند. حتی اگر یکی از این پروتئینها همزمان با بقیه پیدا نشود، تاژک کار نخواهد کرد و انتخاب طبیعی، تاژکی را که کار نمیکند، چرا باید برگزیند؟ مشابه همین استدلال را میتوان برای بال پرندگان به کار برد. شاهپرهای بال تنها در نهایت کمال خود میتوانند به کار پرواز آیند. اما تکامل شاهپرها از کرکپرها ناگهان رخ نداده است. ابتدا پوشپرها تکامل یافتند، بعد شاهپرهای کوتاه با دو نیمه متقارن که به کار پرواز نمیآیند، بعد شاهپرهایی بلندتر تکامل یافتند که همچنان دو نیمه متقارن داشتند و سرانجام شاهپرهایی بلند و محکم پیدا شدند که یک نیمهشان پهنتر از نیمه دیگر بود و این یعنی، شاهپر مورد نظر خاصیت هواپویایی۱۷ هم دارد و میتواند به کار پرواز آید. منتها تا پیش از پیدایش پرواز، انتخاب طبیعی چرا باید شاهپری را برگزیند که به درد پرواز نمیخورد؟
 شکل ۸. حضور شاهپر در دایناسورهای پردار بیپرواز بالغ و فقدان آن دردایناسورهای نابالغ نشاندهنده نقش شاهپر به عنوان صفت ثانویه جنسی است.
شکل ۸. حضور شاهپر در دایناسورهای پردار بیپرواز بالغ و فقدان آن دردایناسورهای نابالغ نشاندهنده نقش شاهپر به عنوان صفت ثانویه جنسی است.
پاسخ این معما در فرایندی تکامل نهفته است به نام همسازگاری.۱۸ همسازگاری نوع خاصی از سازگاری است که در آن صفتی به دلیلی مورد انتخاب طبیعی قرار میگیرد، اما پس از مدتی در کنار چند صفت دیگر، کاربرد جدیدی هم پیدا میکند؛ مثلاً پمپهای پروتونی در غشای یاخته باکتری با انتقال پروتون، ATP میسازند یا مصرف میکنند. ساختار و عملکرد این پمپها با پروتئینهایی که مسئول چرخاندن تاژک هستند، یکی است. این شباهت نشان میدهد پس از آنکه پمپهای پروتونی برای جابهجایی پروتون تکامل یافتند، در کنار پروتئینهایی دیگر (که هرکدام برای استفادههای دیگر تکامل یافته بودند) به کار ساخت تاژک هم آمدند. انتخاب طبیعی تکتک این پروتئینها را پیشتر، برای مقاصد دیگری انتخاب کرده بود؛ نه صرفا برای ساخت تاژک در آینده. شواهد سنگوارهای نشان میدهد شاهپرهای تروپودها نیز برای مقصودی به جز پرواز رشد کردهاند. شاهپرهای بلندی که روی دستها، دم و حتی پاهای دایناسورهای بالغ دیده میشوند، اما در نمونههای نابالغ همین گونههای پردار کوچک یا غایب هستند، نشانه خوبی است که این ساختارها در آغاز صفات ثانویه جنسی بودهاند که برای جفتیابی تکامل یافتهاند. البته این توضیح ضروری است که هنوز هم در پرندگان، شاهپرها کاربرد آغازین خود را حفظ کردهاند و پرندگان زیادی هستند که با شاهپرهای گشوده و حرکاتی موزون، توجه جنس مخالف را جلب میکنند. در بسیاری از پرندگان (مثلاً درناها) هر دو جنس ظاهر یکسان دارند و هر دو پرنده نر و ماده با رقصهای جفتیابی، طرف مقابل را برای تولیدمثل ارزیابی میکنند. بنابراین، نباید انتظار داشت صفات ثانویه جنسی لزوماً در یکی از دو جنس پررنگتر باشد.
پرواز از روی درخت یا از روی زمین؟یکی دیگر از معماهای بزرگ درباره تکامل پرواز این است که نخستین دایناسورهای پردار چگونه پرواز کردند؟ از روی درخت به زمین پریدند یا از روی زمین پر زدند و پریدند؟ اهمیت این سوال وقتی مشخص میشود که بدانیم بیشتر تروپودهای پردار، دایناسورهایی شکارچی و دونده با جثه بوقلمون تا شترمرغ بودهاند. این دایناسورها گرچه شاهپرهای بلندی داشتند، مثل پرندگان بیپرواز یا کمپرواز امروزی، بیشتر روی زمین میدویدند و به صورت گروهی به شکار دایناسورهای بزرگتر میرفتند، اما معدودی از آنها جثههایی کوچکتر داشتند و ساختار دست/بال آنها نشان میدهد دستکم توانایی سر خوردن در هوا به کمک بالهای گشوده را داشتهاند. آرکئوپتریکس یکی از همین دایناسورهای کوچکجثه پردار بود که میتوانست با بالهای گشوده روی هوا سر بخورد.
نه آرکئوپتریکس و نه هیچکدام از نمونههای مشابهی که در اواخر دوره ژوراسیک (۱٤٥ میلیون سال پیش) تا اوایل دوره کرتاسه (۱۲۰ میلیون سال پیش) میزیستند، توانایی بالزدن نداشتند. از سویی استخوان جناغ (Sternum، استخوانی بزرگ که در پرندههای امروزی زیر قفسه قرار دارد و ماهیچههای پروازی سینهای به آن متصل میشود) در آرکئوپتریکس و سایر پرندگان قدیمی بسیار کوچک، ضعیف و حتی غیراستخوانی (غضروفی) است. استخوان جناغ کوچک، یعنی ماهیچههای سینهای کوچک و کوچکبودن ماهیچههای سینهای، یعنی قدرت پایین آمدن بال/دست بسیار ضعیفتر از پرندگان امروزی بوده است.
از سوی دیگر مفصل بازو و کتف به شکلی است که دست/بال نمیتواند به سمت پشت بدن خم شود و حداکثر به سمت طرفین باز میشود؛ بنابراین، هنگام بال زدن، از طرفین (حالت افقی) به سمت پایین بدن (عمودی) حرکت میکرده و بیشترین نیرو در حالی وارد میشده که بالها در پایین بدن به حالت عمودی درآمده بودند. در پرندگان امروزی، دست/بال کاملاً به سمت پشت بدن خم میشود و هنگام بالزدن از حالت عمودی (به سمت بالا) به سمت پایین میآید و بیشترین نیرو زمانی وارد میشود که بالها در دو طرف بدن باز هستند. تغییر سمت و سوی مفصل بازو به سمت پشت مدتها پس از آرکئوپتریکس رخ داد بنابراین، میتوانیم نتیجه بگیریم آرکئوپتریکس قادر به بالزدن نبوده و نخستین پروازها، محدود به سر خوردن در هوا با بالهای گشوده بوده که احتمالاً از شاخهای به شاخه دیگر انجام میشده است.
 شکل ۹: وجود پر در دایناسورها بیش از آنکه تا ۳۰ سال پیش تصور میشد،عمومیت دارد. در این شکل گلهای Zhuchengtyrannus (زوچنگوتیرانوس) جواندیده میشوند که به نبرد خانوادهای Ischioceratops (ایسکیوسراتوپس)رفتهاند.
شکل ۹: وجود پر در دایناسورها بیش از آنکه تا ۳۰ سال پیش تصور میشد،عمومیت دارد. در این شکل گلهای Zhuchengtyrannus (زوچنگوتیرانوس) جواندیده میشوند که به نبرد خانوادهای Ischioceratops (ایسکیوسراتوپس)رفتهاند.
زوچنگوتیرانوس خویشاوند آسیایی تیرانوسورِس است و ایسکیوسراتوپس،سردهای از تبار دایناسورهای شاخدار. سنگوارههای نمایندگان هر دو گروهنشان میدهد این دایناسورها پردار بودهاند. پرهای دایناسورهای شاخدارمومانند و تیغمانند بودند و پرهای دایناسورهای گوشتخوار، بیشتر به پرپرندگان شبیه بود (تصویرسازی امیرحسین بهجتی).
دایناسورها چند بار پرواز کردند؟این فرض که آرکئوپتریکس نیای پرندگان امروزی است، بسیار شایع، اما نادرست است. آرکئوپتریکس ویژگیهایی دارد که نشان میدهد نیای مستقیم پرندگان امروزی نیست. در حقیقت آرکئوپتریکس یکی از چندین گونه دایناسورهای پرداری است که زمانی در کره زمین پراکنده بودند و نیای پرندگان نیز یکی از همین دایناسورها بوده است، اما آرکئوپتریکس با نیای پرندگان تفاوتهایی داشته است. آخرین بررسیهای فیلوژنتیک۲۰ به روشنی نشان میدهد آرکئوپتریکس خویشاوندی نزدیکتری با دایناسورهای شکارچی و پردار مشهوری مثل ولاسیراپتور۲۱ دارد تا با پرندگان امروزی. علاوه بر آرکئوپتریکس، چند نمونه دیگر از دایناسورهای پردار نیز توانایی محدودی در پرواز یا سر خوردن روی هوا داشتهاند. اغلب این نمونهها از جمله راهونهویس۲۲ و مایکروراپتور۲۳ خویشاوندی نزدیکتری با ولاسیراپتور و سایر شکارچیان دونده دوره کرتاسه داشتهاند، اما نکته عجیب اینجاست که این چند دایناسور پروازگر، هرکدام متعلق به تباری هستند که اغلب نمونههای آنها قادر به همین حد از پرواز هم نبودهاند. به نظر میرسد توانایی پرواز در نیای مشترک این دایناسورها وجود نداشته، یا بسیار محدود بوده است.۲۴ گروه دیگری نیز که به پرندگان امروزی منتهی میشوند، همچنین شامل انواعی اولیه هستند که اثری از توانایی پرواز یا سر خوردن در هوا میان آنها دیده نمیشود. بنابراین به نظر میرسد توانایی پرواز در دایناسورهای پردار، دستکم چند بار جداگانه تکامل یافته است. منتها یکی از این گروهها موفقشده است در رقابت با سایر دایناسورهای پردار، پیروز شود و نسل همین گروه که امروزه پرندگان نامیده میشود، تا امروز باقی مانده است.
جالبترین نکته درباره این حقیقت که پرواز چندین بار در دایناسورهای پردار تکامل یافته، این است که همه گروههایی که توانایی پرواز اولیه داشتند، از سازوکاری یکسان برای پریدن استفاده نمیکردند. اسکانسوریوپتریجیدها۲۵ گروهی عجیب از تبار دایناسورهای پردار بودند که گویی به طور ثانویه شاهپرهای پروازی خود را از دست داده بودند. جثه آنها به اندازه کبوتر یا حتی کوچکتر و قدمت آنها (۱٦٥-۱٥٦ میلیون سال پیش) حتی از آرکئوپتریکس هم بیشتر بود. دو سرده بهتر شناختهشده از این خانواده، یی۲۶ و آمبوپتریکس۲۷ به وضوح شواهدی از تکامل نوع متفاوتی بال را نشان میدهند. بال این دایناسورها شبیه بال تروسورها و خفاشها بدون پر و کاملاً پوستی بوده، منتها این پوست میان انگشتان دوم و سوم دست و پهلوها کشیده شده بود. به علاوه، استخوانی دراز و میلهمانند نیز از مچ دست این دایناسورها به سمت عقب کشیده شده بود و پرده پوستی بال را پشتیبانی میکرد. تحلیلهای فیلوژنتیک نشان میدهد این دایناسورها نسبت نزدیکتری با پرندگان امروزی دارند تا آرکئوپتریکس و ولاسیراپتور و سایر دایناسورهای پردار، اما همین که سازوکار متفاوتی برای پرواز در آنها تکامل یافته است، نشان میدهد تکامل پرواز چندین بار به صورتهای مختلف در دایناسورها رخ داده است و تبار پرندگان، گروهی از دایناسورهای پردار بودهاند که از آغاز توانایی پرواز نداشتهاند و بعدها بهتدریج و در مسیری جدا از آرکئوپتریکس، این توانایی در پرندگان تکامل و توسعه یافته است.
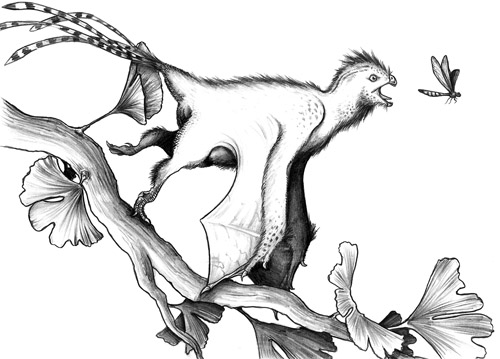
شکل ۱۰: یی ( Yi) دایناسوری از خویشاوندان پرندگان بود که با سازوکاریمتفاوت پرواز میکرد.
بالهای پوستی و خفاشمانند این دایناسور نشان میدهداستعداد پرواز در بسیاری تروپودها وجود داشته، اما تجلی آن به صورت پرواز،بارها به صورت جداگانه و با سازوکارهایی متفاوت رخ داده است (تصویرسازیامیرحسین بهجتی).
خزندگانی با استعداد پروازاگر امروز تروسورها زنده بودند، به احتمال زیاد در فرهنگ عامه به عنوان گروهی از مرغان شناخته میشدند. بررسیهای دیرینهشناختی و تکاملی نشان میدهد بسیاری از ویژگیهایی که پرندگان را مستعد تکامل پرواز کرد، از قبیل سوخت و ساز خونگرم، وجود پر و کیسههای هوایی متصل به ششها در میان تروسورها و پرندگان خاستگاه مشترکی دارند. علاوه بر این میدانیم که نه یک گروه، بلکه دستکم پنج گروه مختلف از دایناسورهای پردار، تا حدودی توانایی پریدن و سر خوردن در هوا و پرواز را به دست آورده بودند. پرندگان تنها بازماندگان از دایناسورهای پردار هستند. تمام اینها نشان میدهد فرضیه همسازگاری چقدر درست و دقیق است. سازگاریهای منتهی به پرواز در همه این خزندگان به دلایلی دیگر تکامل یافته است، ولی بعدها در پرواز هم نقش بازی کردند؛ مثلاً، کیسههای هوایی احتمالاً برای بهبود کارایی تنفس و افزایش سوختوساز در نیای مشترک دایناسورها و تروسورها تکامل یافتند. این کیسهها به دایناسورهای بزرگجثه کمک کردند جثههایی بسیار غولآسا پیدا کنند، اما با جرم بسیار کمتر از چیزی که انتظار میرفت. اگر جرم حجمی دایناسورهای غولآسا به اندازه پستانداران میبود، جرم بزرگترین دایناسورها باید به حدود ۱٥۰ تن میرسید، اما در حقیقت جرم این دایناسورها حداکثر بین ٥۰ تا ۸۰ تن بوده است و این یعنی فشار مکانیکی کمتر به ماهیچهها و استخوانها به لطف بدنی که پر از کیسههای هوایی است. همین کیسههای هوایی بدن تروپودهای پردار و نمونههای امروزی آنها، یعنی پرندگان را نیز سبکتر و نقش مهمی در تکامل پرواز ایفا کردند. اگر پرندهای به جثه گربه (مثلاً مرغ خانگی) را در یک دست بگیرید و گربهای را در دست دیگر، متوجه خواهید شد که بدن پرنده چقدر سبکتر از پستاندار همجثه است. باقی صفات منتهی به تکامل پرواز نیز به همین صورت باعث شدند که گروههای متعددی از تروپودهای پردار، مستعد پریدن میان شاخ و برگ درختان باشند. این نکته تکاملی شاید برای گروههای دیگر جانداران نیز صادق باشد که تکامل صفات شاخص برخی گروهها، شاید چندان آنی و دفعی نبوده باشد و سازگاریهای اولیه، مدتها پیشتر در نیاکان گروه موردنظر پیدا شده باشند یا صفت مورد نظر، مدتها بعد از تکامل آن گروه با به هم پیوستن سازگاریهای مختلف، «کامل شده» باشد.
پینوشتها
1. Archaeopteryx
2.Thomas Henry Huxley
3. در مقالههایی در سال ۱۸٦۸، سالنامه و نشریه تاریخ طبیعی به نشانی goo.gl/khD6aZ، صفحه ٦٦ و ۱۸۷۰، فصلنامه انجمن زمینشناسی به نشانی goo.gl/KnkCYM
4. در سال ۱۸۸٤ اوتنیل چارلز مارش، دیرینهشناس و مکتشف بزرگ آمریکایی نیز که بسیاری از دایناسورهای مشهور آمریکا را در غرب وحشی کشف کرده بود، ردهبندی مفصلی از دایناسورهای کشفشده تا آن زمان ارائه کرد و به شباهت آرکیوپتریکس و یکی از دایناسورهای شکارچی بهنام سراتوسورِس (Ceratosaurus)که خودش پیدا کرده بود (نیچر به نشانی goo.gl/itBEBX) اشاره کرد. مارش هم نتیجه گرفت که آرکیوپتریکس و پرندهها دایناسور هستند. دیرینهشناس مشهور دیگری که نظر هاکسلی را دنبال کرد فرانتز نوپچا، دیرینهشناس اشرافزاده مجار بود که در سال ۱۹۰۷ مقالهای درباره تکامل پرواز نوشت (شرح اقدامات انجمن جانورشناسی لندن به نشانی bit.ly/ 205 Bsot) و در آن نتیجه گرفت آرکیوپتریکس از نسل دایناسورهای دونده (و نه درختزی) تکامل یافته است.
5. ریچارد اُون، توصیفکننده نخستین سنگواره آرکیوپتریکس، مشهور به نمونه [موزه تاریخ طبیعی] لندن، هم مخالف سرسخت نظریه تکامل داروین بود و طبعاً مخالف تکامل پرندهها از دایناسورها. مخالفت او باعث شد انعکاس خبر کشف سنگواره آرکیوپتریکس در برخی کتابهای تاریخ طبیعی بهخصوص در فرانسه بدون کوچکترین دلالتهای تکاملی باشد.
6. Gerhard Heilmann
7. به دانمارکی Vor Nuvaerende Viden om Fuglenes Afstamning و ترجمه انگلیسی آن The Origin of Birds
8. Clavicle
9. هنری فرفیلد آزبورن ولاسیراپتور (Velociraptor)، سورونیتوئیدس (Saurornithoides) و اُویراپتور (Oviraptor) را در مقالهای در تجربیات موزه تاریخ طبیعی آمریکا توصیف کرده که هرسه از خویشاوندان نزدیک پرندگان به شمار میروند. در صفحه ۱۱ این مقاله تصویر اسکلت اُویراپتور دیده میشود و ترقوه پرندهمانند آن با علامت Ic (مخفف Interclavicle یا میانترقوه) مشخص شده است. مقاله را در این نشانی خواهید یافت: bit.ly/ 2Ddbwp
10. جان اُستروم با کشف داینونیکوس (Deinonychus) در ۱۹٦٤ و توصیف آن در ۱۹٦۹ (خبرنامه موزه تاریخطبیعی پیبادی، goo.gl/t8T33G) و توصیف نمونه جدیدی از آرکیوپتریکس در ۱۹۷۰ (ساینس، bit.ly/ 2XJCALW) شباهتهای زیادی میان داینونیکوس -که دایناسوری شکارچی و دونده به جثه گرگ بود- و آرکیوپتریکس نشان داد و طی سالهای بعد این نظریه را پیگیری کرد (۱۹۷۳، نیچر: ۱۹۷٥، سالنامه مروری علوم زمین و سیارهای: bit.ly/2qwQM5P، ۱۹۷٦، نشریه زیستشناختی انجمن لینهای: bit.ly/35y2SKM). رابرت باکر و پیتر گالتون در سال ۱۹۷٤ مقالهای انقلابی منتشر کردند و نشان دادند نه تنها پرندهها از دایناسورها مشتق شدند، بلکه آنها را جزء دایناسورها ردهبندی کردند (نیچر: goo.gl/eM68Ay). باکر همین ادعای انقلابی را در مقالهای در سال ۱۹۷٥ (ساینتیفیک آمریکن: bit.ly/2L6DYDZ) و در کتاب مهم و تأثیرگذاری تکرار کرد که در سال ۱۹۸٦ با عنوان عقاید نامتعارفی درباره دایناسورها (bit.ly/37DJXCL) منتشر شد.
11. Jacques Gauthier
12. مقاله گوتیه در کتاب خاستگاه پرندگان و تکامل پرواز منتشر شد؛ قابل بازیابی در: bit.ly/2KjpBT1
13. برای اطلاعات بیشتر درباره مبانی ردهبندی پرندگان به عنوان زیرگروهی از خزندگان رک: عرفان خسروی، «آیا پرندگان خزندهاند؟»، رشد آموزش زیستشناسی ۲۹:۲، زمستان ۱۳۹۲، صص ٦-۱٦؛ به نشانی:9640 / roshdmag.ir/fa/article
14.Archosauromorpha
15. Lepidosauromorpha
16. Ornithischian Dinosaurs
17. Aerodynamic
18. Exaptation
19. Adaptation
20. اسکات هارتمن و همکارانش اخیراً جامعترین و دقیقترین تحلیل تبارشناختی درباره تکامل دایناسورهای پردار را در دل مقالهای منتشر کردند که به توصیف گونهای جدید از تروپودهای پرندهمانند از آمریکای شمالی میپردازد. تحلیل تبارشناختی این مقاله عمدتاً حاصل کار بیستساله نویسنده دوم مقاله، میکی مورتیمر است. او از اواخر دهه ۱۹۹۰ مشغول جمعآوری اطلاعات، مقایسه و تصحیح دادههای برآمده از تحلیلهای تبارشناختی تروپودها بوده است. این مقاله در این نشانی قابل دسترس است: 7247/ peerj.com/articles
21.Velociraptor
22. Rahonavis
23.Microraptor
24. البته این نظر مورد توافق همه دیرینهشناسان نیست. هنرمند و دایناسورشناس دیگری بهنام گرگوری پاؤل در کتابی که در سال ۱۹۸۸ بهنام دایناسورهای شکارچی جهان (bit.ly/ 23bolox) منتشرکرد، مدعی شد آرکیوپتریکس نهتنها عموی بزرگ پرندگان امروزی است، بلکه بسیاری از دایناسورهای شکارچی دونده غیردرختزی و غیرپروازگر مشهور ـ که از سوی دیرینهشناسان دیگر، پسرعموهای پرندگان تلقی شده بودند- در حقیقت پرندههایی ابتدایی بودند که از نسل پرندههایی کوچکجثه مثل آرکیوپتریکس تکامل یافته، ولی مثل شترمرغهای امروزی، پرواز را کنار گذاشته و بهطور ثانویه بیپرواز شدهاند. او در تمامی کتابها و آثار بعدی خود نیز از همین نظریه دفاع کرده است.
25. Scansoriopterygidae
26. Yi
27. Ambopteryx